Что такое биосинтез днк
Содержание:
- Роль комплементарности в механизме матричного синтеза
- Ферменты
- ДНК (дезоксирибонуклеиновая кислота)
- Репликация (редупликация) ДНК
- Строение и функции РНК
- Что представляют собой белки
- Дезоксирибонуклеиновая кислота: коротко о главном
- Репликация ДНК и ее строение
- Генетический код и его свойства
- Влажная B-ДНК
- Трансляция
- Особенности репликационного комплекса
- Схема биосинтеза белка
- Суперскрученная молекула
Роль комплементарности в механизме матричного синтеза
Принцип комплементарности основан на выборочном соответствии азотистых оснований нуклеотидов друг другу. Так, аденину в качестве пары подойдут только тимин или урацил (двойная связь), а гуанину — цитозин (3 тройная связь).

В процессе синтеза нуклеиновых кислот со звеньями одноцепочечной матрицы связываются комплементарные нуклеотиды, выстраиваясь в определенную последовательность. Таким образом, на основании участка ДНК ААЦГТТ при репликации может получиться только ТТГЦАА, а при транскрипции — УУГЦАА.
Как уже было отмечено выше, белковый синтез происходит с участием посредника. Эту роль выполняет транспортная РНК, которая имеет участок для присоединения аминокислоты и нуклеотидный триплет (антикодон), предназначенный для связывания с матричной РНК.
В этом случае комплементарный подбор происходит не по одному, а по три нуклеотида. Так как каждая аминокислота специфична только к одному виду тРНК, а антикодон соответствует конкретному триплету в РНК, синтезируется белок с определенной последовательностью звеньев, которая заложена в геноме.
Ферменты
В процессе репликации участвует большое количество различных ферментов. Каждый из них несет особую функцию в процессе.
ДНК – полимераза
ДНК – полимераза – это фермент, ответственный за создание ДНК из нуклеотидов, точнее строительных блоков для ДНК. ДНК – полимераза должна копировать молекулы двухцепочечной ДНК. Как раз именно этот процесс называется принципом полуконсервативности. Во время деления клетки ДНК занимается дублированием ее же самой. Так копия исходной молекулы передается к каждой из дочерних клеток вместе с генетической информацией.
Иногда же ДНК – полимераза может ошибиться, обычно это повторяется через каждый миллиард скопированных пар оснований. Поэтому данный фермент отвечает еще и за корректировку нити ДНК.
Также ДНК – полимераза может добавлять новые и свободные нуклеотиды к 3’ – концу, тем самым образуя новые нити. Однако ДНК – полимераза не может самостоятельно начинать новую цепь.
ДНК — лигаза
ДНК – лигаза – фермент, которые соединяет разрывы в отстающей цепочке ДНК. Лигаза образует фосфодиэфирные связи между свободными 3’ и 5’ концами. Для образования фосфодиэфирной связи ДНК – лигаза использует энергию, полученную из гидролиза (АТФ).
В 1961 году два американских генетика, а именно М. Мезельсон и Д. Вейгл, поняли, что при рекомбинации происходит разрыв и соединение ДНК. Это дало толчок ученым к поиску фермента, который мог бы сшивать разделенные фрагменты ДНК. В 1967 году М. Мезельсон и Д. Вейгл находят нужный фермент и называют его ДНК – лигазой.
ДНК – геликаза (хеликаза)
ДНК – геликаза – фермент, который раскручивает двухцепочечную спираль ДНК. Раскручивая цепи, данный фермент разделяет нити между собой, делая их одинарными. ДНК – геликаза движется по одноцепочечной нити, как только он встречает участок с двумя закрученными цепями, он разрывает водородные связи между основаниями, тем самым продвигая репликативную вилку дальше
ДНК – праймаза
Праймаза – фермент, необходимый для инициации репликации ДНК. Этот фермент синтезирует ДНК – праймеры, которые и запускают синтез матричной цепи. Также ДНК – праймеры запускают синтез фрагментов Оказаки для запаздывающей цепи.
ДНК – топоизомераза
ДНК – топоизомераза – фермент, который изменяет степень сверхспиральности2. Из-за суперспиральности образуется напряжение в спирали, которое в итоге мешает репликации. Чтобы такого не случалось, существует такой фермент как ДНК – топоизомераза.
Белки SSB
SSB – это белки, которые помогают сохранить нити ДНК в состоянии, когда они расплетены, а также они соединяют одноцепочечные фрагменты ДНК. Такие белки предотвращают комплементарное спаривание.
ДНК (дезоксирибонуклеиновая кислота)
ДНК (дезоксирибонуклеиновая кислота) – своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение.
ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни человека.
Дезоксирибонуклеиновая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
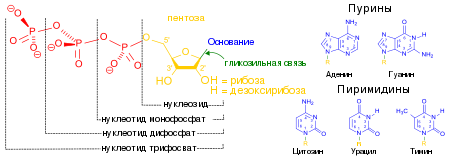
ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы.
Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи).
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей.
Вторичная структура ДНК представляет собой двойную спираль, состоящую из двух параллельных неразветвленных полинуклеотидных цепей, закрученных в противоположные стороны вокруг общей оси.
Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы – снаружи.
Две спирали удерживаются вместе водородными связями между парами азотистых оснований. Водородные связи образуются между определенными основаниями: тимин (Т) образует водородные связи только с аденином (А), а цитозин (Ц) – только с гуанином (Г). В первой паре азотистых оснований две водородные связи, а во второй – три.
Такие пары оснований называются комплементарными парами. А такое пространственное соответствие молекул, способствующее их сближению и образованию водородных связей, называется комплементарностью. Комплементарность обусловливает спиралевидную модель ДНК.
Две спирали в молекуле ДНК комплементарны друг другу. Последовательность нуклеотидов в одной из спиралей определяет последовательность нуклеотидов в другой.
В каждой паре оснований, связанных водородными связями, одно из оснований – пуриновое, а другое пиримидиновое. Общее число остатков пуриновых оснований в молекуле ДНК равно числу остатков пиримидиновых оснований.
Таким образом,
- ТИМИН (Т) комплементарен АДЕНИНУ (А),
- ЦИТОЗИН (Ц) комплементарен ГУАНИНУ (Г).
Комплементарность полинуклеотидных цепей служит химической основой главной функции ДНК – хранения и передачи наследственных признаков.
Репликация ДНК
Двухспиральная структура ДНК с комплементарными полинуклеотидными цепями обеспечивает возможность самоудвоения (репликации) этой молекулы.
Перед удвоением водородные связи разрываются, и две цепи раскручиваются и расходятся. Каждая цепь затем служит матрицей для образования на ней комплементарной цепи.
После разделения цепей происходит саморепликация, т.е. образование новой двойной спирали, идентичной исходной.
После репликации образуются две дочерние молекулы ДНК, в каждой из которых одна спираль взята из родительской ДНК, а другая (комплементарная) синтезирована заново.
Таким образом, сохраняется и передается новому поколению исходная структура ДНК.
Длина полинуклеотидных цепей ДНК практически неограничена. Число пар оснований в двойной спирали может меняться от нескольких тысяч у простейших вирусов до сотен миллионов у человека.
Видеофильм «ДНК. Код Жизни»
Рубрики: Нуклеиновые кислоты
Репликация (редупликация) ДНК
Репликация ДНК — процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным.
«Строительным материалом» и источником энергии для репликации являются дезоксирибонуклеозидтрифосфаты (АТФ, ТТФ, ГТФ, ЦТФ), содержащие три остатка фосфорной кислоты. При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка фосфорной кислоты отщепляются, и освободившаяся энергия используется на образование фосфодиэфирной связи между нуклеотидами.
В репликации участвуют следующие ферменты:
- геликазы («расплетают» ДНК);
- дестабилизирующие белки;
- ДНК-топоизомеразы (разрезают ДНК);
- ДНК-полимеразы (подбирают дезоксирибонуклеозидтрифосфаты и комплементарно присоединяют их к матричной цепи ДНК);
- РНК-праймазы (образуют РНК-затравки, праймеры);
- ДНК-лигазы (сшивают фрагменты ДНК).
С помощью геликаз в определенных участках ДНК расплетается, одноцепочечные участки ДНК связываются дестабилизирующими белками, образуется репликационная вилка. При расхождении 10 пар нуклеотидов (один виток спирали) молекула ДНК должна совершить полный оборот вокруг своей оси. Чтобы предотвратить это вращение ДНК-топоизомераза разрезает одну цепь ДНК, что дает ей возможность вращаться вокруг второй цепи.
ДНК-полимераза может присоединять нуклеотид только к 3′-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3′-конца к 5′-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по-разному и в противоположных направлениях. На цепи 3’–5′ синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей. На цепи 5’–3′ — прерывисто, фрагментами (фрагменты Оказаки), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстающей).
Купить проверочные работы по биологии
Особенностью ДНК-полимеразы является то, что она может начинать свою работу только с «затравки» (праймера). Роль «затравок» выполняют короткие последовательности РНК, образуемые при участи фермента РНК-праймазы и спаренные с матричной ДНК. РНК-затравки после окончания сборки полинуклеотидных цепочек удаляются.
Репликация протекает сходно у прокариот и эукариот. Скорость синтеза ДНК у прокариот на порядок выше (1000 нуклеотидов в секунду), чем у эукариот (100 нуклеотидов в секунду). Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3′-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Что представляют собой белки
Белок является важнейшим функциональным элементом в клетках всех живых организмов. Они выполняют каталитическую, структурную, регуляторную, сигнальную, защитную и множество других функций.
Молекула белка – это биополимер, образованный последовательностью аминокислотных остатков. Она, как и молекулы нуклеиновых кислот, характеризуется наличием нескольких уровней структурной организации – от первичного до четвертичного.
Существует 20 видов обладающих определенными особенностями (канонических) аминокислот, используемых живыми системами для построения огромного количества самых разнообразных белков. Самостоятельно белок, как правило, не синтезируется. Ведущая роль в формировании сложной белковой молекулы принадлежит нуклеиновым кислотам – ДНК и РНК.
Дезоксирибонуклеиновая кислота: коротко о главном
ДНК — как карта памяти, в ней вся информация, необходимая для развития любого организма. Как именно проявится эта информация, во многом зависит от того, как работают другие нуклеиновые кислоты — РНК. Но про них расскажем в другой раз.
Структура ДНК известна всем: знаменитую двойную спираль постоянно показывают в фантастических фильмах про мутантов и используют в качестве иллюстрации к научпоп-статьям.

Каждая спираль состоит из нуклеозидов, их может быть четыре типа:
- Аденозин (А)
- Тимидин (Т)
- Гуанозин (Г)
- Цитидин (Ц)
Это те буквы, которыми написан текст жизни. Каждое слово в нем — отдельный ген. Каждый в какой-то степени определяет наше здоровье, способности, внешний вид — практически все.
Чтобы жизнь на Земле продолжалась, клетки (а вместе с ними и ДНК) должны делиться. Необходимость в делении клеток возникает также при росте организма и обновлении тканей. При этом ДНК клетки сначала удваивается, а затем распределяется по новым клеткам.
Репликация ДНК и ее строение
И первичная, и вторичная структуры ДНК возникают в ходе явления, называемого репликацией – образования из материнской макромолекулы двух идентичных ей «двойных спиралей». При репликации исходная молекула расплетается, и на освободившихся одиночных цепочках происходит наращивание комплементарных оснований. Поскольку половинки ДНК антипараллельны, этот процесс протекает на них в разных направлениях: по отношению к материнским цепочкам от 3’-конца к 5’-концу, то есть новые цепочки растут в направлении 5’ → 3’. Лидирующая цепь синтезируется непрерывно в сторону репликационной вилки; на отстающей цепи синтез совершается от вилки отдельными участками (фрагменты Оказаки), которые затем сшивает между собой особый фермент – ДНК-лигаза.
Пока продолжается синтез, уже сформированные концы дочерних молекул претерпевают винтообразное закручивание. Затем, еще до окончания репликации новорожденные молекулы начинают образовывать третичную структуру в процессе, именуемом сверхспирализацией.
Генетический код и его свойства
Генетический код — система записи информации о последовательности аминокислот в полипептиде последовательностью нуклеотидов ДНК или РНК. В настоящее время эта система записи считается расшифрованной.
Свойства генетического кода:
- триплетность: каждая аминокислота кодируется сочетанием из трех нуклеотидов (триплетом, кодоном);
- однозначность (специфичность): триплет соответствует только одной аминокислоте;
- вырожденность (избыточность): аминокислоты могут кодироваться несколькими (до шести) кодонами;
- универсальность: система кодирования аминокислот одинакова у всех организмов Земли;
- неперекрываемость: последовательность нуклеотидов имеет рамку считывания по 3 нуклеотида, один и тот же нуклеотид не может быть в составе двух триплетов;
- из 64 кодовых триплетов 61 — кодирующие, кодируют аминокислоты, а 3 — бессмысленные (в РНК — УАА, УГА, УАГ), не кодируют аминокислоты. Они называются кодонами-терминаторами, поскольку блокируют синтез полипептида во время трансляции. Кроме того, есть кодон-инициатор (в РНК — АУГ), с которого трансляция начинается.
Таблица генетического кода
| Первоеоснование | Второе основание | Третьеоснование | |||
|---|---|---|---|---|---|
| У(А) | Ц(Г) | А(Т) | Г(Ц) | ||
| У(А) | ФенФенЛейЛей | СерСерСерСер | ТирТир—— | ЦисЦис—Три | У(А)Ц(Г)А(Т)Г(Ц) |
| Ц(Г) | ЛейЛейЛейЛей | ПроПроПроПро | ГисГисГлнГлн | АргАргАргАрг | У(А)Ц(Г)А(Т)Г(Ц) |
| А(Т) | ИлеИлеИлеМет | ТреТреТреТре | АснАснЛизЛиз | СерСерАргАрг | У(А)Ц(Г)А(Т)Г(Ц) |
| Г(Ц) | ВалВалВалВал | АлаАлаАлаАла | АспАспГлуГлу | ГлиГлиГлиГли | У(А)Ц(Г)А(Т)Г(Ц) |
* Первый нуклеотид в триплете — один из четырех левого вертикального ряда, второй — один из верхнего горизонтального ряда, третий — из правого вертикального.
Влажная B-ДНК
При малом содержании солей и высокой степени гидратации, то есть в нормальных физиологических условиях, ДНК принимает свою главную форму B. Природные молекулы существуют, как правило, в В-форме. Именно она лежит в основе классической модели Уотсона-Крика и чаще всего изображается на иллюстрациях.
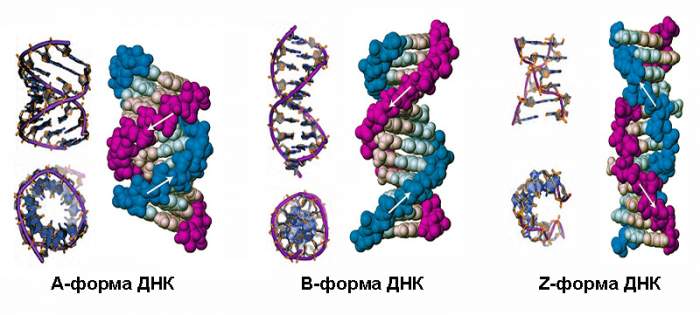
Данной форме (она также правозакрученная) свойственна меньшая компактность размещения нуклеотидов (0,33 нм) и большой шаг винта (3,3 нм). Один виток содержит 10,5 пары оснований, поворот каждой из них относительно предыдущей составляет около 36°. Плоскости пар почти перпендикулярны оси «двойной спирали». Диаметр такой сдвоенной цепочки меньше, чем у А-формы – он достигает только 2 нм.
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, — рибосомы. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три — в пептидильном и три — в аминоацильном участках.
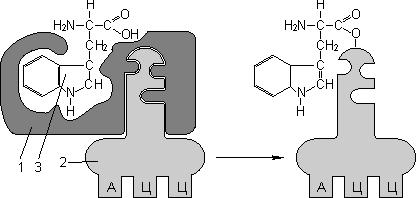
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК (лекция №4). Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3′-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.
 |
||
|
1 — антикодон;2 — участок, связывающий аминокислоту. |
Транспорт аминокислот к рибосомам: 1 — фермент; 2 — тРНК; 3 — аминокислота. |
Двадцать видов аминокислот кодируются 61 кодоном, теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.
Синтез белка начинается с того момента, когда к 5′-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислот.
Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
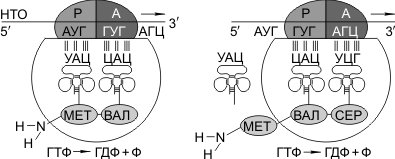
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе в течение трех лет принимали участие 10 человек.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Особенности репликационного комплекса
Репликация протекает также с участием сложной совокупности ферментов – реплисомы, главную роль в которой играет ДНК-полимераза.

Одна из цепочек в ходе биосинтеза ДНК является лидирующей и формируется непрерывно. Образование отстающей цепи проходит путем присоединения коротких последовательностей – фрагментов Оказаки. Эти фрагменты сшиваются при помощи ДНК-лигазы. Такое течение процесса называется полунепрерывным. Кроме того, его характеризуют как полуконсервативный, поскольку в каждой из новообразованных молекул одна из цепей – материнская, а вторая – дочерняя.
Репликация ДНК – это один из ключевых этапов при клеточном делении. Данный процесс лежит в основе передачи наследственной информации новому поколению, а также роста организма.
Схема биосинтеза белка
По схеме видно, как протекает процесс.
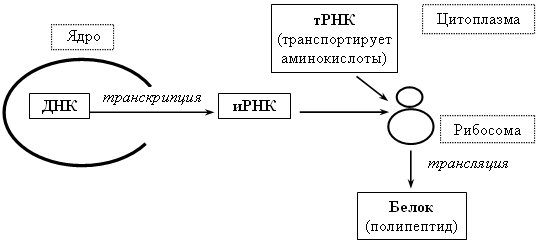
Точкой стыковки этой схемы являются рибосомы, в которых синтезируется белок. В простой форме синтез осуществляется по схеме
ДНК >, PHK >, белок.
Первым начинается этап транскрипции, в котором молекула изменяется в одноцепочную информационную рибонуклеиновую кислоту (иРНК). В ней содержится информация аминокислотной последовательности белка.
Следующей остановкой иРНК будет рибосома, в которой происходит сам синтез. Происходит это путём трансляции, формирования полипептидной цепочки. После этой заурядной схемы, полученный белок транспортируется в разные места, выполняя определённые задачи.
Суперскрученная молекула
Сверхспирализованная форма ДНК возникает, когда двухцепочечная молекула совершает дополнительное закручивание. Оно может быть направлено по часовой стрелке (положительно) либо против (в этом случае говорят об отрицательной сверхспирализации). ДНК большинства организмов суперскручена отрицательно, то есть против основных витков «двойной спирали».
В результате образования дополнительных петель – супервитков – ДНК приобретает сложную пространственную конфигурацию. В клетках эукариот этот процесс происходит с формированием комплексов, в которых ДНК отрицательно навивается на гистоновые белковые комплексы и принимает вид нити с бусинами-нуклеосомами. Свободные участки нити называются линкерами. В поддержании суперскрученной формы молекулы ДНК принимают участие и негистоновые белки, а также неорганические соединения. Так образуется хроматин – вещество хромосом.
Хроматиновые нити с нуклеосомными бусинами способны к дальнейшему усложнению морфологии в процессе, называемом конденсацией хроматина.