Строение животной клетки
Содержание:
- Аппарат Гольджи
- Физико-химические свойства мембран
- Поры
- Формирование и рост клеточных оболочек
- Плазмодесмы
- Химический состав и строение плазматической мембраны
- Ослизнение клеточных оболочек
- Клеточный цикл
- Осмотическое давление в клетке
- Механизмы проникновения веществ в клетку
- Функции клеточной стенки
- Плазмодесмы
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Физико-химические свойства мембран
При обычных (свойственных организму) условиях физическое состояние мембран — жидкое. Однако в их молекулярной организации есть порядок (представлен выше), поэтому правильно называть ее жидкокристаллическим, а состояние этой кристаллической структуры — смектическое.
Свойства:
- чувствительность к внешним условиям;
- асимметричность;
- текучесть;
- изменчивость;
- самоорганизация;
- замкнутость;
- пластичность.
Особенности этой организации, свойственные цитолеммам, позволяют им перейти и в другое состояние (например, в гель при понижении температуры).
Именно поэтому при длительном изменении внешних условий в мембранах происходит изменение и химического состава — они проходят период адаптации, что не всегда благотворно сказывается на состоянии клетки.
При химическом анализе установлено, что все элементы, входящие в состав клеточной оболочки по количеству вариативны. Например, в эритроцитах количество белковых молекул в 2,5 раза больше, чем липидных, а в миелиновой мембране — наоборот.
Поры
Внутренне утолщение клеточной стенки не бывает вполне равномерным. Сформировавшаяся оболочка имеет более толстые и менее утолщенные участки. Даже в тех случаях, когда стенки, в общем, имеют равномерную толщину, в них, при детальном рассмотрении обнаруживаются небольшие углубления. Эти места, в которых оболочка очень тонка, и называются порами.
Таким образом, поры у растений — это не сквозные многочисленные отверстия, как это понимается в общеупотребительном смысле. У растений порой называют любое неутолщенное место оболочки.
Для обозначения сквозных отверстий у растений используется другое название — перфорации.
Поры в 2-х сосединх клетках располагаются одна против другой, образуя так называемую пару пор.
У клеток с мощной вторичной оболочкой поры в разрезе имеют вид радиальных каналов. На поперечном срезе эти каналы могут иметь разную форму: чаще округлую, реже щелевидную (эллиптическую или крестообразную). Округлые поры обычно формируются в паренхимных клетках, щелевидные — в прозенхимных. По форме порового канала обычно различают поры 2-х типов: простые и окаймлённые.
Простые поры имеют достаточно ровный канал, с одинаковым диаметром на всём протяжении.
У окаймлённых пор голосеменных растений на первичной оболочке образуется линзовидное утолщение — торус, а выросты вторичных оболочек как бы нависают над торусом.
Окаймлённые поры характерны для водопроводящих элементов древесины. Эти элементы имеют вид длинных труб разного диаметра. По этим трубкам, как по капиллярам, поднимается вода. Понятно, что давление воды в смежных клетках неодинаково. В этом случае торус смещается и прижимается к выступам вторичной оболочки клеток с меньшим давлением.
Обычно к порам приурочены и плазмодесменные канальцы. Нередко через одну пору проходят десятки плазмодесм.
В любом случае, поры, как и плазмодесмы, облегчают диффузию веществ, растворённых в воде, из одной клетки в другую.
Сквозные отверстия клеточных стенок — перфорации особенно характерны для водопроводящих поперечных перегородок водопроводящих члеников сосудов. Как правило, в этих перегородках образуются одна, две или несколько крупных перфораций.
Многочисленные мелкие перфорации имеются в так называемых ситовидных трубках, по этим трубкам также передвигается вода с органическими веществами, но сверху вниз, от листьев к корням.
Формирование и рост клеточных оболочек
Даже молодые меристематические клетки снабжены первичной оболочкой. Новая оболочка формируется в процессе деления клетки в заключительной стадии митоза — телофазе. Заключительная в телофазе стадия — процесс деления цитоплазмы на дочерние клетки называется цитокинезом.
В ранней телофазе между двумя дочерними ядрами формируется бочкообразная система волокон, называемая фрагмопластом. Волокна фрагмопласта, так же как и волокна митотического веретена состоят из микротрубочек. В световом микроскопе видно, как в экваториальной плоскости фрагмопласта появляются мелкие капли, которые затем сливаются, образуя клеточную пластинку. Клеточная пластинка растёт центробежно до тех пор, пока не достигнет оболочки делящейся клетки. С помощью электронного микроскопа было установлено, что сливающиеся капельки — это пузырьки, отрывающиеся от аппарата Гольджи. Они в основном содержат пектиновые вещества, из которых формируется срединная пластинка, а мембраны пузырьков участвуют в построении плазматической мембраны по обеим сторонам пластинки. В это время из фрагментов трубчатого эндоплазматического ретикулума образуются плазмодесмы.
После образования срединной пластинки протопласт дочерних клеток откладывает на нее первичную оболочку.
Слой целлюлозы, который откладывается во время роста клетки, называется первичной клеточной оболочкой. Помимо целлюлозы, гемицеллюлоз и пектина первичные оболочки содержат и структурный белок — гликопротеин. Первичные оболочки могут и лигнифицироваться, хотя, как правило, лигнин им не свойственен. Однако наиболее характерную часть первичной оболочки составляет пектиновый компонент. Он придаёт оболочке пластичность, позволяет ей растягиваться, по мере удлинения органов: корня, стебля, листа.
Пектиновые вещества способны сильно набухать, поэтому первичные оболочки содержат много воды (60-90%).
В целом, на долю гемицеллюлоз и пектиновых веществ, приходится 50-60 % сухого веса первичной оболочки, содержание целлюлозы не превышает 30 %, структурный белок занимает до 10%, лигнин, как правило, отсутствует.
Вторичное утолщение оболочек обычно начинается после прекращения роста клеток. При этом толщина оболочки увеличивается, а объём, запасаемый полостью клетки, сокращается.
Во многих случаях по завершении вторичного утолщения протопласты клеток отмирают, но клетки продолжают функционировать, выполняя главным образом механическую и опорную функцию.
В соответствие с механической функцией строение и химический состав вторичной оболочки сильно отличается от первичной. В ней значительно меньше воды и преобладают плотно сомкнутые микрофибриллы целлюлозы (40-50 %), в первичной оболочке — они расположены рыхло. Много во вторичных оболочках и лигнина — 25-30%, он придаёт оболочкам дополнительную жёсткость и прочность; гемицеллюлозы составлют 20-30% и практически нет пектиновых веществ.
Итак: лицо первичной оболочки составляют пектиновые вещества, в то время как вторичной — лигнин.
Вторичная оболочка далеко не всегда откладывается равномерно. У некоторых специализированных водопроводящих клеток она имеет вид колец или спиральных лент. Такие клетки сохраняют способность к продольному растяжению и после отмирания.
Плазмодесмы
Протопласты соседних клеток связаны между собой тонкими нитями цитоплазмы — плазмодесмами. Эти структуры присущи только растительным клеткам.
В нормальном состоянии плазмодесмы невидимы в световой микроскоп, однако, если стимулировать набухание оболочки плазмодесмы, становятся заметными, поэтому выявлены и описаны они были уже достаточно давно. Хотя детали строения плазмодесм изучены сравнительно недавно с помощью электронного микроскопа. Под электронным микроскопом плазмодесмы выглядят как узкие каналы (диаметром от 30 до 60 нм), выстланные плазматической мембраной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера — десмотрубочка, которая сообщается с эндоплазматическим ретикулом обеих смежных клеток.
Десмотрубка напоминает цитоплазматические микротрубочки или жгутики простейших. Она состоит из 11 спирально расположенных белковых субъедениц.
Вокруг десмотрубки локализируется цитоплазма, которая во многих типах плазмодесм непосредственно не соединяется с цитоплазмой клеток.
В плазмодесмах обнаружена АТФ — азная активность.
Наличие плазмодесм обеспечивает непрерывность цитоплазмы клеток, составляющих органы и ткани. Такая непрерывная система называется симпласт. Кроме того, за счёт плазмодесм обеспечивается единство эндоплазматической сети, переходящей из клетки в клетку. Единая эндоплазматическая сеть получила название эндопласт.
Таким образом, выделяется три непрерывных компартмента в растительных тканях — это:
- единая цитоплазма — симпласт,
- непрерывный ретикулум — эндопласт и соприкасающиеся между собой клеточные стенки, вместе с межклетниками составляют непрерывную систему — апопласт.
Размещаются плазмодесмы в стенке либо группами, либо равномерно разбросаны по всей стенке.
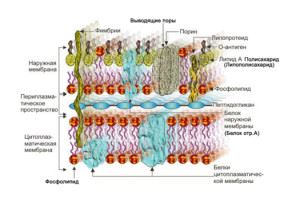
Химический состав и строение плазматической мембраны
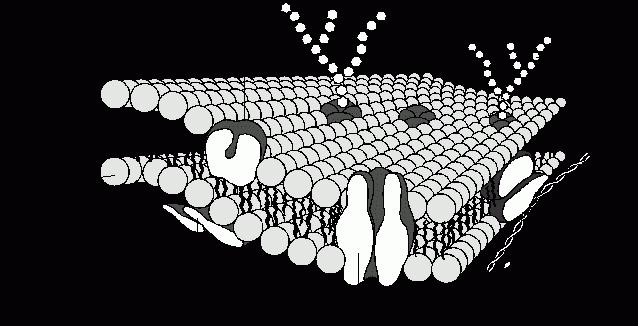
Плазматическая мембрана, окружая каждую клетку, отделяет ее содержимое от внеклеточного пространства. В состав мембраны входят липиды, белки и углеводы. Основой плазмалеммы является двойной слой из фосфолипидов. Молекула фосфолипида имеет небольшую гидрофильную «головку» (остатки глицерина, ортофосфорная кислота и дополнительные соединения) и два (реже один) длинных гидрофобных «хвоста» (остатки жирных кислот). Гидрофобные части молекул объединяются с другими гидрофобными соединениями, а гидрофильные — с гидрофильными, формируя двойные слои, как показано на рисунке.
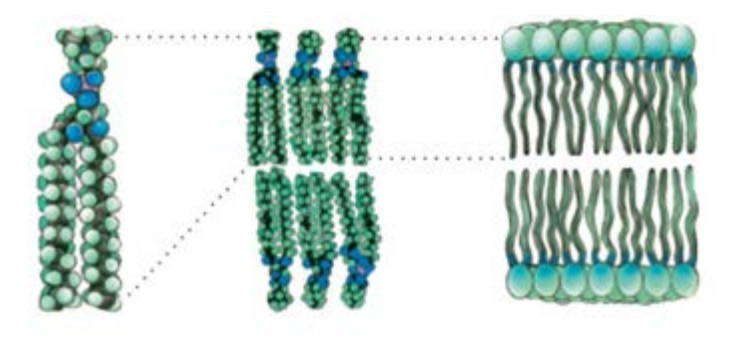
Схема расположения фосфолипидов в мембране клетки
В каждом слое гидрофильные «головки» молекул обращены к водной среде (внеклеточное пространство или цитоплазма), а их «хвосты» ориентированы внутрь толще мембраны. Такую структуру имеют все биологические мембраны, в том числе и внутриклеточных органелл.
Кроме фосфолипидов в состав плазматической мембраны входят другие липиды (в частности, холестерол) и значительное количество белков (до 50% от массы мембраны). Поскольку белковые молекулы по размеру больше, чем фосфолипидные, на один белок в составе мембраны приходится около 50 фосфолипидов. В зависимости от функций клетки количество и состав мембранных белков существенно различаются. По расположению в мембране разделяют белки, пронизывающие толщу мембраны (внутренние или интегральные) и такие, которые размещены с внутренней или внешней стороны мембраны (внешние или периферийные). Мембранные белки могут соединяться с углеводами (вспомните, как они называются) как на иллюстрации ниже.

Схема строения плазматической мембраны
Такая модель строения биологических мембран получила название жидкостно-мозаичной: большинство липидов мембраны находятся в жидком состоянии и лишь около 30% липидов прочно соединены с внутренними белками в комплексные соединения.
С плазматическими мембранами связан надмембранный комплекс — набор структур, расположенных снаружи клеток.
Надмембранный комплекс животных клеток представляет собой углеводороды части гликопротеинов и гликолипидов мембран, образующих наружный слой клетки — гликокаликс, который выполняет рецепторную и маркерную функции, а также участвует в обеспечении избирательности транспорта веществ и пристеночном (примембранном) пищеварении.
У бактерий, растений и грибов надмембранный комплекс представлен клеточной стенкой — жестким каркасом, окружающим клетки. Клеточные стенки разных организмов имеют разную химическую природу. Вы уже знаете, что основным веществом стенок растительных клеток является целлюлоза. У грибов эта структура сформирована другим полисахаридом — хитином. Бактериальные клетки окружены стенками из пептидогликана (также известный как муреин) — вещества сложной химической природы (содержит короткие пептиды и остатки углеводов).
Ослизнение клеточных оболочек
При ослизнении клеточных оболочек образуются слизи и камеди. Те и другие представляют собой высокомолекулярные углеводы, состоящие большей частью из пентоз и их производных. Они нерастворимы в спирте, эфире, а в воде сильно набухают.
Резкой границы между ними не установлено. Обычно их различают по консистенции в набухшем состоянии: камеди клейки и могут вытягиваться в нити, слизи же сильно расплываются и в нити не тянутся.
В сухом состоянии камеди и слизи очень тверды и хрупки, и лишь при смачивании водой они переходят в тягучее желеобразное состояние.
Ослизение бывает или нормальным явлением в жизни растения, или же происходит как патологический процесс.
В некоторых случаях слизи и камеди появляются в клеточных оболочках путём отложения на ранее образовавшиеся слои клеток, иногда они образуются из имеющегося уже материала стенок в результате химического метаморфоза — «слизистого перерождения».
При патологическом ослизнении — камедетечении (или гуммозе) — в камедь превращаются и стенки, и содержание клеток; процесс этот захватывает большие участки тканей. Гуммоз часто наблюдается у вишен, слив, персиков, у многих акаций и астрагалов.
Значение ослизнения клеточных стенок во многих случаях очевидно. Например, ослизненные наружные слои клеток кожицы семян, набухая весной, входят в соприкосновение с почвой. Слизь, благодаря клейкости, закрепляет семена на влажном месте и, поглощая воду из почвы, улучшает водный режим проростка, передавая ему воду и защищая от высыхания.
Слизь может использоваться как запасное, питательное вещество.
Клеточный цикл
Согласно научным источникам, в клеточный цикл входят все периоды развития клетки от момента деления материнской и образования дочерней до гибели (или деления). Клеточный цикл кратко можно охарактеризовать несколькими точными параметрами.
Длительность
Существуют как быстро делящиеся — 12-36 ч (например, кроветворные), так и медленно воспроизводящиеся. Средний цикл, свойственный многим организмам — от 10 до 25 часов.
Фазы клеточного цикла
Жизнь клеточного организма можно разделить на несколько фаз.
Фазы:
- Интерфаза, или клеточный рост. В этот период происходит быстрая наработка веществ (ДНК, белков и т. д.) и подготовка к делению. Интерфазу можно условно разделить на подпериоды. Это G1-фаза (начальный рост), S-фаза (репликация ДНК) и G2-фаза (непосредственно подготовка к митозу).
- Фаза митоза, или фаза М. Это время жизни также можно разделить на две стадии – кариокинез (деление ядра) и цитокинез (деление цитоплазмы).
Клеточный цикл — высокоорганизованная система.
Регуляция клеточного цикла
Все периоды клеточного цикла регулируются особыми белками — циклин-зависимыми киназами и циклинами. Содержание этих белков варьируется на разных стадиях жизненного цикла. После митотического деления они полностью разрушаются.
Осмотическое давление в клетке
Осмотическое давление — избыточное гидростатическое давление на раствор, отделённый от чистого растворителя полупроницаемой мембраной, при котором прекращается диффузия растворителя через мембрану (осмос). Это давление стремится уравнять концентрации обоих растворов вследствие встречной диффузии молекул растворённого вещества и растворителя.
Мера градиента осмотического давления, то есть различия водного потенциала двух растворов, разделённых полупроницаемой мембраной, называется тоничностью. Раствор, имеющий более высокое осмотическое давление по сравнению с другим раствором, называется гипертоническим, имеющий более низкое — гипотоническим.
Тургор тканей — напряжённое состояние оболочек живых клеток. Тургорное давление — внутреннее давление, которое развивается в растительной клетке, когда в неё в результате осмоса входит вода и цитоплазма прижимается к клеточной стенке; это давление препятствует дальнейшему проникновению воды в клетку.
Тургор обуславливается тремя факторами: внутренним осмотическим давлением клетки, которое вызывает напряжение клеточной оболочки, внешним осмотическим давлением, а также упругостью клеточной оболочки.
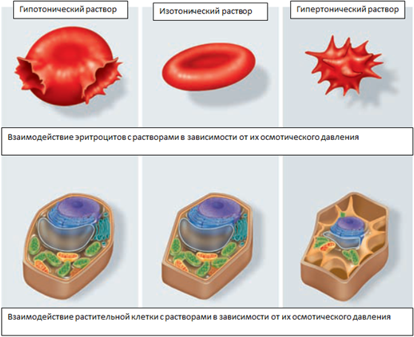
Рисунок 11. Взаимодействие эритроцитов и растительной клетки с растворами.
Механизмы проникновения веществ в клетку
Между клеткой и окружающей средой постоянно происходит обмен веществ. Ионы и небольшие молекулы транспортируются через мембрану путем пассивного или активного транспорта, макромолекулы и крупные частицы — путем эндо- и экзоцитоза.
| Способ переноса | Направление переноса | Переносимые вещества | Затраты энергии | Описание способа |
|---|---|---|---|---|
| Диффузия: через липидный слой (пассивный транспорт) | По градиенту концентрации | O2, CO2, мочевина, этанол | Без затрат энергии (пассивный процесс) | Мелкие нейтральные молекулы просачиваются между молекулами липидов. Гидрофобные вещества, как правило, диффундируют быстрее гидрофильных. Ионы и крупные молекулы не могут пересечь липидный бислой |
| Диффузия: через белковые поры (пассивный транспорт) | Ионы (в том числе Ca2+, K+, Na+), вода | Трансмембранные (интегральные) белки могут иметь водные каналы, по которым ионы или полярные молекулы пересекают мембрану, минуя гидрофобные хвосты липидов | ||
| Облегченная диффузия (пассивный транспорт) | Глюкоза, лактоза, аминокислоты, нуклеотиды, глицерин | Белок-переносчик, находящийся в клеточной мембране, на одной стороне мембраны присоединяет молекулу или ион. Это изменяет форму молекулы переносчика, и его положение в мембране изменяется так, что молекула или ион выделяются уже с другой стороны мембраны | ||
| Активный транспорт | Против градиента концентрации | Na+ и K+, H+, аминокислоты в кишечнике, Ca2+ в мышцах, Na+ и глюкоза в почках | С затратами энергии (активный процесс) | Как и облегченная диффузия, осуществляется белками-переносчиками. Но в данном случае изменение формы молекулы переносчика (ее конформация) вызывается присоединением не молекулы переносимого вещества, а фосфатной группы, отделившейся от молекулы АТФ в ходе гидролиза. |
| Фагоцитоз | Крупные макромолекулы и твердые частицы | В месте контакта с частицами мембрана впячивается, затем формируется пузырек, который отшнуровывается от плазматической мембраны и поступает в цитоплазму. Характерен для амебоидных простейших, кишечнополостных, клеток крови — лейкоцитов, клеток капилляров костного мозга, селезенки, печени, надпочечников | ||
| Пиноцитоз | Капли жидкости | Поглощение капель жидкости по механизму, аналогичному фагоцитозу. Характерен для амебоидных простейших и клеток крови — лейкоцитов, клеток печени, некоторых клеток почек |
Пассивный транспорт — перемещение веществ по градиенту концентрации; осуществляется без затрат энергии путем простой диффузии, осмоса или облегченной диффузии с помощью белков-переносчиков.
Диффузия — транспорт ионов и молекул через мембрану из области с высокой в область с низкой их концентрацией, т.е. по градиенту концентрации. Диффузия может быть простой и облегченной. Если вещества хорошо растворимы в жирах, то они проникают в клетку путем простой диффузии. Например, кислород, потребляемый клетками при дыхании, и углекислый газ в растворе быстро диффундируют через мембраны. Вода способна проходить также через мембранные поры, образованные белками, и переносить молекулы и ионы растворенных в ней веществ.
Осмос — диффузия воды через полупроницаемую мембрану из области с меньшей концентрацией солей в область с более высокой их концентрацией. Возникающее давление на полупроницаемую мембрану называют осмотическим. Клетки содержат растворы солей и других веществ, что создает определенное осмотическое давление. Живые клетки способны регулировать его, изменяя концентрацию веществ. Например, амебы имеют сократительные вакуоли для регуляции осмоса. В организме человека осмотическое давление регулируется системой органов выделения.
Облегченная диффузия — транспорт веществ в клетку через ионные каналы, образованные в мембране белками, с помощью белков-переносчиков, также находящихся в мембране. Таким образом попадают в клетку нерастворимые в жирах и не проходящие через поры вещества. Например, путем облегченной диффузии глюкоза поступает в эритроциты.
Активный транспорт — перенос веществ белками-переносчиками против градиента концентрации с затратами энергии. Например, транспорт аминокислот, глюкозы, ионов натрия, калия, кальция и др.
Эндоцитоз — поглощение веществ (путем окружения) выростами плазматической мембраны с образованием окруженных мембраной пузырьков. Экзоцитоз — выделение веществ из клетки (путем окружения) выростами плазматической мембраны с образованием окруженных мембраной пузырьков. Поглощение и выделение твердых и крупных частиц получило названия фагоцитоз и обратный фагоцитоз, жидких или растворенных частичек — пиноцитоз и обратный пиноцитоз соответственно.
Функции клеточной стенки
Оболочки разных клеток совместно обеспечивают всему растению и его отдельным частям механическую прочность и опору. Это функция клеточной стенки аналогична одной из функций скелета животных. Однако она не единственная.
Жесткость стенок препятствует растяжению клеток и их разрыву. В результате по физическим законам в клетки может путем осмоса поступать вода. Для травянистых растений тургоцентричность клеток является единственной их опорой.
Микрофибриллы целлюлозы ограничивают рост клеток и определяют их форму. Если микрофибриллы окольцовывают клетку, то она будет расти в длину (поперек направления волокон).
Связанные клеточные стенки образуют апопласт, по которому передвигается вода и минеральные вещества. Плазмодесмы связывают содержимое разных клеток в единую систему — симпласт.
Стенки сосудов ксилемы, трахеид, ситовидных трубок выполняют транспортную функцию.
Наружные клеточные стенки эпидермальных клеток покрыты воском (кутикулой). С одной стороны, он препятствует испарению воды, с другой – проникновению вредных микроорганизмов.
У некоторых растений в определенных клетках оболочки видоизменяются и служат местом запаса питательных веществ.
Плазмодесмы
Протопласты соседних клеток связаны между собой тонкими нитями цитоплазмы — плазмодесмами. Эти структуры присущи только растительным клеткам.
В нормальном состоянии плазмодесмы невидимы в световой микроскоп, однако, если стимулировать набухание оболочки плазмодесмы, становятся заметными, поэтому выявлены и описаны они были уже достаточно давно. Хотя детали строения плазмодесм изучены сравнительно недавно с помощью электронного микроскопа. Под электронным микроскопом плазмодесмы выглядят как узкие каналы (диаметром от 30 до 60 нм), выстланные плазматической мембраной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера — десмотрубочка, которая сообщается с эндоплазматическим ретикулом обеих смежных клеток.
Десмотрубка напоминает цитоплазматические микротрубочки или жгутики простейших. Она состоит из 11 спирально расположенных белковых субъедениц.
Вокруг десмотрубки локализируется цитоплазма, которая во многих типах плазмодесм непосредственно не соединяется с цитоплазмой клеток.
В плазмодесмах обнаружена АТФ — азная активность.
Наличие плазмодесм обеспечивает непрерывность цитоплазмы клеток, составляющих органы и ткани. Такая непрерывная система называется симпласт. Кроме того, за счёт плазмодесм обеспечивается единство эндоплазматической сети, переходящей из клетки в клетку. Единая эндоплазматическая сеть получила название эндопласт.
Таким образом, выделяется три непрерывных компартмента в растительных тканях — это:
- единая цитоплазма — симпласт,
- непрерывный ретикулум — эндопласт и соприкасающиеся между собой клеточные стенки, вместе с межклетниками составляют непрерывную систему — апопласт.
Размещаются плазмодесмы в стенке либо группами, либо равномерно разбросаны по всей стенке.